안녕하세요?
지난번 시간까지 선천적 or 내재적 면역 반응(innate immune response)에 대해서 이야기를 했다면, 이번 포스팅부터 한동안은 적응면역(adaptive immunity)에 대해서 포스팅을 하고자 합니다. 우선 가장 기본이 되는 항원과 항체에 대한 이야기를 하기 전에, 지난번 까지 줄기차게 이야기를 했던 선천적 or 내재적 면역계와 무엇이 다른지에 대해서 간략하게 소개를 해 보겠습니다.
먼저 인체는 피부와 점막 등으로 일차적인 방어선을 형성합니다. 그리고 나서 이 일차적인 방어선을 뚫고 미생물이 침입하면, 이차적인 방어선으로 선천적 or 내재적 면역이 미생물을 지난 포스팅에서 이야기한 것처럼 제거를 시작합니다. 이 단계에서도 침입한 미생물을 제거하지 못하면, 이제 세번째 방어 시스템인 이 적응면역(adaptive immunity)가 활동해서 침입자를 제거하려는 과정을 시도합니다.

위 그림은 3중 성벽으로 유명한 콘스탄티노플의 3중 성벽입니다. 우리 몸의 방어체계도 3개의 성벽으로 쌓인 것처럼 '물리적 방어막', '내재적 혹은 선천적 면역' 그리고 마지막으로 '적응면역'이 병원체의 침입으로 부터 인체를 보호하고 있는 것 입니다. 그럼 3중 성벽의 마지막 벽에 해당하는 적응면역반응은 병원체를 인식하는 단계에서 부터 앞서 소개가 된 선천적 면역 반응과는 다른 전략을 사용합니다.
먼저 지난번 포스팅에서 '조혈모세포'에서 부터 림프계 전구체가 나오고, 이 림프계 전구체로 부터 B cell, T cell이 나온다는 것을 그림으로 묘사를 하였습니다.
링크 : 선천적 or 내재적 면역 이야기 보충 - 선천적 면역과 적응 면역의 연결고리
이 B cell과 T cell은 각각 '골수'와 '흉선'에서 분화하기 때문에 이름이 이렇게 붙은 것인데, 중요한 것은 이 B cell과 T cell에는 각각 '세포 수용체(cell receptor)'라는 것이 존재하며, 거의 무한대로 다양한 형태로 수백만개 이상이 B cell과 Tcell의 세포 표면에 존재하고 있습니다.

뜬금없이 왠 항체그림이냐고 하실 분이 많은데, 항체란 것도 별것아닌 것이, 먼저 B cell의 세포 수용체는 다른 이름으로 '면역글로불린 (immunoglobulin)'이라고도 불린다고 합니다. 다중에 다루겠지만, B cell이 더 분화하게 되면 이 '면역글로불린'을 자기 세포막의 표면만이 아니라 인체의 '체액'에도 녹는 면역글로불린을 분비한다고 합니다. 바로 이 '용해성 면역글로불린'이 다른 말로는 '항체(antibody)'라고 불리우는 것입니다.
결국 우리가 '항체'라고 부르는 것이 실제로는 B cell과 T cell의 표면에도 있고, 혈액 속에도 항체가 자리잡고 있다고 합니다. 그리고 이 '항체'에 인식이 되는-결합이 가능한 거대분자, 바이러스 및 박테리아, 기생충, 단백질, 이식 된 장기 및 심지어는 우리몸의 일부분(자가항원)을 '항원(antigen)'이라고 부르게 되는 것입니다. 당연 그 종류에는 제한이 있을까 하는 의문이 들 정도로 다양합니다.

위 그림은 항체의 중요한 특징을 며사한 그림입니다. 우선 위 그림에서 홍역 바이러스에 '특이성(specificity)'이라는 것이 있는데, 말 그대로 '항원' 이되는 '홍역바이러스'에는 특이성이 있는 상체들이 반응을 합니다. 하지만 그림에서 우측에 그려진 항체들은 '홍역 바이러스'에만 '특이성'이 있어서 반응을 하지, 다른 '인플루엔자 바이러스'에는 마치 자물쇠에 '안맞는' 열쇠처럼 결합을 하지 못합니다.

먼저 '항체'가 되는 면역글로불린의 구조에 대해서 간단하게 소개를 할까 합니다. '중쇄(heavy chain)'와 '경쇄(light chain)'이라고 불리는 2개의 서로 다른 폴리 펩타이드로 이루어져 있으며, Y자 형상의 모양을 하고 있습니다. 이 Y자의 끝 부분을 '가변부위(variable region)'이라고 하는데, 이 부분은 직접 항원과 '결합'하는 부위이며, 다양한 항원이 있듯이 가변부위 역시 셀수 없이 많은 변형이 있습니다.
반면에 '일정부위(constant region)'는 항체마다 아미노산의 서열이 언제나 같은 부위입니다. 그럼 이러한 항체가 언제나 Y자 형태만 가지느냐 하면 그것도 아닌게, IgM, IgA, IgE, IgD, IgG로 구조가 나누어져 있습니다. 다만 이번 포스팅에서는 여기까지는 다루지 않도록 하겠습니다. 다만 마지막으로 이 항체가 선천적 면역 반응에서 언급이 되었는 보체의 활성화에 어떻게 관여하는 지를 설명하겠습니다.

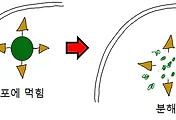
먼저 C1 복합체(C1 component)라는 단백질이 여기서 등장을 하게 됩니다. 이 C1은 항체가 박테리아에 붙은 다음에, 아래의 그림에서 묘사된 것과 같이 항체와 결합을 하게 됩니다.

이 단계에서 부터 보체의 '고전 경로(classical pathway)'가 시작되는 것입니다. 먼저 일어나는 일은 C1이 구조적으로 '비활성화'된 상태에서 '활성화'되는 상태로 변하는 것입니다. 그러면 '또 다른' 보체인 C4를 C4a와 C4b로 자르게 됩니다.

여기서 '또 다른' 보체인 C2가 등장하게 되는데, 재미있게도 C2는 C4b와 합체해서 C4b2라는 단백질을 만들어 내게 되고, 이 C4b2가 또 활성화된 C1에 의해서 잘리는 것입니다. 즉, C1은 활성을 듸고서 C4를 '두번 자르는 것'입니다.

여기서 생성된 C4b2a가 이전 포스팅에서 언급을 한 것처럼 C3라는 보체를 C3a와 C3b로 잘라서 면역반응을 유도합니다.
링크 : 선천적 or 내재적 면역 이야기 part2-보체 이야기
다시 말하지만, 보체의 '고전경로(classical pathway)'가 이번 포스팅에서 간략학 소개가 되었으며, 이후의 C3가 잘린 후로는 '대체경로(alternative pathway)'와 같은 길을 걷게 됩니다.
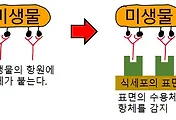
조금은 헷갈릴 수도 있지만, 선천적 면역 반응에서 '대체경로'로 보체가 활성화 되었다면, 적응면역에서는 '고전경로'라고 해서, 병원체에 항체가 붙는 것으로 보체가 활성화 된다고 보시면 됩니다. 다만 이 보체의 활성 이외에도 적응 면역과 관계된 것은 많기에 이만 줄이며, 다음 포스팅에서는 항체가 식세포(대식세포나 호중구)의 활동을 도우며, 어떻게 항체가 생성되는 지에 대해서 올리겠습니다.
'생명과학 이야기 > 면역학 이야기' 카테고리의 다른 글
| 적응면역 이야기 part3-T cell 이야기 part1 (0) | 2017.08.12 |
|---|---|
| 적응 면역 이야기 part2-항체의 생성 이야기 (0) | 2017.08.11 |
| 선천적 or 내재적 면역 이야기 보충 - 선천적 면역과 적응면역의 연결고리 (0) | 2017.08.05 |
| 선천적 or 내재적 면역 이야기 final-선천적 면역의 마지막 이야기 (0) | 2017.08.01 |
| 선천적 or 내재적 면역 이야기 part3-보체의 이후 이야기 (0) | 2017.07.31 |